
Ever since French beekeepers saw their bees dying as they collected pollen from treated sunflowers back in 1996, beekeepers have been concerned that their bees are being harmed the highly toxic neonicotinoid insecticides, with imidacloprid most widely used. The use of this class of insecticide has grown steadily ever since. Bee losses have become chronic as well. However, unlike the first case in France where bees were literally falling dead while gathering pollen, the widespread colony losses today are less explainable, often associated with outbreaks of a variety of diseases, and with very high winter colony mortality. So why blame the insecticides?
To see why the bees are dying, and why these pesticides are still being sold, we must examine the toxicology of the neonicotinoid chemicals as well as the history and science of pesticide regulation. The toxic nature of a chemical is characterized by its “LD50” level. This is the amount of chemical that will kill half of the test organisms in short order. For many traditional pesticides (organophosphates), the LD50 level provides an adequate overall characterization of toxic effect. These pesticides tend to be short-lived and generally do not bio-accumulate in the target organism. If the dose doesn’t kill the organism, the toxic compound will be metabolized and excreted. Since the organophospate pesticides — which for many years made up the majority of pesticides sold — could be characterized easily with the single LD50 number, the culture of pesticide regulation largely accepts the acute LD50 as determinative for all toxic effects.
The acute LD50 characterization works poorly for substances that bio-accumulate and/or have a relatively long time-to-effect characteristic. Substances that fall into this category are heavy metals that are known to accumulate in certain tissues, and some carcinogens where an initial single exposure can give rise to cancers much later in the life of the organism. Neonicotinoid insecticides also fit into this category. These insecticide molecules bond strongly and irreversibly at nicotine receptor sites in the central nervous system. There is also evidence of delayed time-to-effect of several days for exposures below the acute LD50 [Suchail et. al.(2001)].
Toxicologists attempt to model the time-dependent effects of chemicals at various dose levels [Tennekes, (2010)]. One of the simplest empirical models assumes the dose/effect relationship can be characterized by a simple “power law” where the effect is proportional to the dose multiplied by the time-of-exposure raised to a power, b.
Effect = Dose x Timeb
Instead of a single number, now we have two numbers to characterize toxicity, the dose and the time exponent. When a power law is plotted with logarithmic scales, one gets a straight line with slope equal to the exponent, b, and intercept equal to the Dose. The time-independent, acute LD50 model is the special case when b=0. For a simple bio-accumulation model, one would expect linearity in time with b=1. Time-to-effect mechanisms require b>0. Combinations of effects for a given organism and chemical will result in a Dose vs Effect curve with characteristic slope that includes all of the time-dependent mechanisms embedded in the value of the exponent.
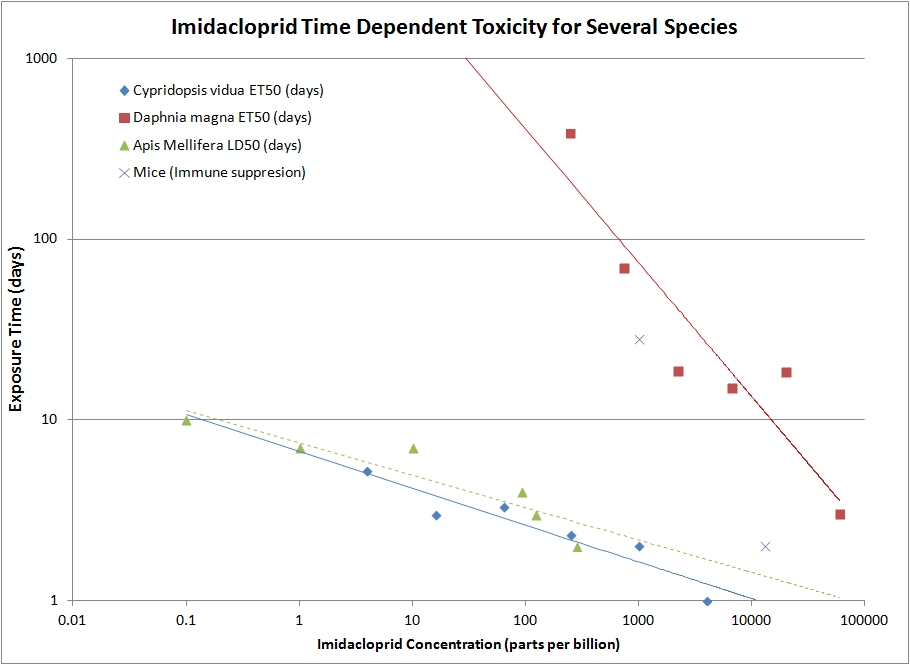
The plot above shows some published data on imidacloprid toxicity on a log-log graph. Data for two species of small aquatic crustaceans, Daphnia magna (red squares) and Cypridopsis vidua (blue diamonds) illustrates the large range of toxicity difference between organisms, and also the large range of applicability of the model, spanning more than three orders of magnitude in concentration. [Sánchez-Bayo et. al. 2009] Honeybees (green triangles) also have high toxicity over a wide range of concentrations [Suchail et al.(2001)].
The best-fit power law curves are shown in the plot. In all cases, the fit parameter, R2, is greater than 0.85. The time exponent for Daphnia, 1.3, is barely more than linear, where as for Cypridopsis and honeybees the exponent is close to 5. This means that there are strong time-dependent delayed effects from the chemical for the Cypriopsis and honeybees. The time5 dependence of the toxicity of imidacloprid for honeybees is the big problem. Very low levels of exposure, with sufficient time, will be lethal. The power law model suggests that we should extend the time of exposure to the lifetime of the organism in order to determine the minimum dose that will have no effect. Honeybees, including the larval stage, live ~50 days in the summer and ~100 days for wintering bees. Extrapolating the green dashed line trying to reach 50 to 100 days would require reducing continuous exposure to less than 0.0001 parts per billion (ppb). This minuscule exposure level is far below the detectable limits of present technology (~1 ppb). What we can detect are residual levels in nectar and pollen on treated plants commonly in the 1 – 10 ppb range; even these small levels are more than 10000 times the level that the power law model suggests would cause no harm to adult bees. Looked at another way, if one bee in 10,000 returns to the colony with pollen gather from a treated plant, that would be enough toxin to begin to cause damage to the colony.
Collecting toxicity data about bees is a complex process, involving multiple trials, caged bees to limit their activity to the test sample, etc. Aquatic crustaceans provide an easier test subject where dosage can be controlled by dilution of the water they live in. The fact that the crustaceans conform to the power law model confirms that imidacloprid is bio-accumulative. The same finding by Suchail et.al. for bees should not be surprising, given that both organisms have central nervous systems with nicotine receptors.
What about mammals and humans? Very little time-dependent information is available. There are a couple of data points on the plot above for mice. One point was the LD50 for short time exposure and the other the threshold for immune system compromise exposed for 28 days. The data for mammals is just not available yet, but even much more modest sensitivity to the chemical could present problems for long-lived species such as humans. There is also fear that these biologically persistent chemicals could be fueling world-wide wild life declines in many species [Mason, R. et. al. 2012].
Our regulators, (EPA) are not adequately considering the time-dependent nature of the lethal effects of the neonicotinoid class of pesticides. The most toxic and long-lived chemicals, imidacloprid, clothanidin, and thiamethoxam should be removed from the market before more harm is done, as is happening in Europe. Other insecticides in this class should be subject to further scrutiny as well.
The social nature of bees naturally draws one to a human analogy. Imagine that a toxic chemical is slowly poisoning our brains. (Think lead pipes and the Romans.) Instead of healthy people living into their 70’s, the toxic effects are bringing on Alzheimer’s-like symptoms to folks in their 40’s and 50’s. The younger healthier part of the population has its hands full, providing for themselves and those that no longer support their own livelihood. Everyone is hungry. Now introduce a bad case of the flu, or the plague, and the already weakened population is devastated. That is what our bees are facing today. The levels of poison are rising as more and more of these pesticides are being used, building up in our soil and in treated plants. The bees are dying younger, and we are gradually eliminating a host of insects and creatures we don’t even know we are poisoning.
Sign our petition to request local stores to stop selling these insecticides.
Sánchez-Bayo, F.,2009. From simple toxicological models to prediction of toxic
effects in time. Ecotoxicology 18,343–354.

Thanks, Gary Reading your discussions of bee poisoning feels like Greek tragedy and moves one to seek change per Aristotle in the POETICS> basically awe, not anger, at seeing through the innocent blindness of human folly. Last week, Scott Meisner told me about willful blindness of the Oregon DEQ in Bend that made me furious. Scott’s son is @ Liz’ age and probably went to South with her. He became an environmental engineer and works out of Portland now. He does a lot of field work and while in Bend saw the Burlington Northern replacing tracks along the Deschutes River–and simply dumping treated ties down the river bank with many jutting into the river. He went directly to the manager of the DEQ office and took a patronizing dismissal of his concern for his trouble. However, Scott’s son also went to South with the fellow who is Kitzhaber’s environmental policy adviser, the one who showed up in the helicopter that afternoon. The next am, BN railroad received a stop work order pending the removal of @ 60,00 (!!!) ties from the Deschutes River watershed! BN legal went nuts and tried their special rights of railroads BS that had intimidated the Bend DEQ. Kitzhaber was not amused, and the clean up is in progress. Despite the happy ending to this drama, the flagrancy of the powerful blinding the watch dogs adds fury to awe at their brazen self serving against the interests of the people. This Bill Moyers discussion with Richard Wolff is somewhat hopeful of a public backlash to scroflaw capitalism, very worth watching: http://billmoyers.com/segment/richard-wolff-on-fighting-for-economic-justice-and-fair-wages/ Charles
Date: Mon, 4 Mar 2013 00:39:33 +0000 To: c_hkittleson@msn.com
Nice story Charles! Chalk up one for the good guys!
Well, we don’t have any proof, that humans_ are NOT gradually being poisoned, by world.- saturation of pesticides. the chances are,we ARE being poisoned, too! The bees are only the first symptoms Our systems become easily vulnerable to poisons, such as, example, auto-exhaust. Researchers say, now, the high increase in breast cancer, is due to auto- exhaust-particles in the air, which react with breast-hormones, to produce cancel cells.
Ask researchers, such as:UCMED CENTER, SAN FRANCISCO, Parnassus Ave.,CA. I saw the results of that research, stating the above findings, years ago. Of course, no one is going to stop driving cars, just because they know the consequences.Being able to have their cars, is more important to everyone, than curing breast cancer in women–and you all KNOW THIS.
I have become a very good judge of human beings, and my prediction is,your pesticide users are NOT GOING TO STOP USING THEM. NO,they will not. Why?Same as the need to drive cars; that;s their priority, and to them, its “To hell with bees,or people, business comes first.”
Just as the Chinese are destroying the air,and China, pesticide users,and corporations do not CARE. they have no scruples.And they are NOT GOING TO STOP producing and using them,unless you FORCE THEM.
THAT is my prediction, and i’m not often wrong about people.
Hi, I live on River Road, a block up from-lrving;; my family made a fruit orchard during the 50’s, we had wonderful fruit for many years, My dad found out DDT was deadly, stopped using it or any pesticides, and we went” organic gardening” from then on,
However now,my plum tree in the front, in the last couple yrs.no longer bears fruit. the roses don’t bloom. I barely get flowering weeds!
the people, here, l talk to, especially very senior, say” the climate here, over several decades, has become so bad, good gardening here is often impossible” they’re right the growing climate here, Now, is TERRIBLE. I had moldy-squash; one July was so cold and damp! I never see any bees here,, where years ago, plenty of bumble-bees,bumbled around us. Fruit trees get destroyed by Spring storms.You need GREEN-HOUSES here, now to garden!
Yes,probably all Willamette Valley is infected with deadly pesticides;the Californians who came here,and others here probably have pesticided the region into “Death.Valley” .Once the bees vanish,we will be next. the bees are only the canarys in the coal mines; We also absorb pesticides. this could easily be the beginning of our end,also.
Nice to hear from a neighbor! Despite your observations that the climate is worse here today than it once was, this region is much better off than a good fraction of the country. Unfortunately, as climate change really kicks in, we will get even more Californians, and Arizonans, and New Mexicans, etc. where lack of water will make those places totally uninhabitable. So… we got to keep this place CLEAN! Most people don’t WANT to foul their nest, they just don’t know they are doing it. Life ultimately is a tragedy, but in the mean time we do what we can.
Hey Gary, You may be well aware of the Xerces Society based in Portland, but just in case, I’m attaching the link: http://www.xerces.org/neonicotinoids-and-bees/ The program director whose umbrella includes the bees/bumbles is Sarina Jepsen. She is from Eugene. A friend of mine, Al Johnson, says she is a fearless advocate and skilled schmoozer–starting way back as a Marist student who sided with the City on the Willamette bike path against the Marist administration and Board. Her bug science ed was at UC Davis. I was just thinking Xerces might offer a link for your research into a serious public policy org. Chas
Date: Mon, 4 Mar 2013 00:39:33 +0000 To: c_hkittleson@msn.com
Thanks Charles, I’ve sent a couple of e-mails into the void with no response from them, so a specific person, Eugene, connection, etc. is good. I agree, we need to work with Xerces.
GR
Gary- I stumbled onto your data, a mere 5 years after you wrote it. I am a scientist, but these days I’m partly-retired and I help people with organic cultivation. I was trying to make an assessment of the risks associated with imidacloprid. NO SURPRISE – our country really hasn’t got good data. It is somewhat disturbing that researchers are saying this chemical binds permanently to neuroreceptors. What would be helpful .. more data related to cancer. Is there anything new in 2018? Thanks, Pete in California.
Hi Pete, I haven’t been following the issue as closely as I was a few years ago, but the problems associated with the neonics have not gone away. I’m not aware of cancer concerns, but what is new are a couple of studies explicitly looking at immune system suppression with these chemicals. Turns out that the same nicotinoide receptor that is found on nervous system synapses is also found on some immune system cells. Here is a new article by G. DiPrisco: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5647381/ . There are probably many other — check the paper bibliography. To me, this mechanism explains much of the devastation we have seen with these chemicals. Immune suppression leaves you with weakened creatures that succumb to diseases and parasites – allowing plausible deniability for the chemical companies since the chemical itself is not the killer. The EU has acted to further restrict the use of these chemicals, but now in the time of Trump, there is no hope that anything will happen at the Federal level in the foreseeable future.